97年度第2學期生命科學系
【遺傳學】課程大綱
教師:李
炎
使用學分/時數:3/3
發講議為課本(印發1份,由同學自行影印):主要參考資料為:
Phillip E. McClean
http://www.ndsu.nodak.edu/instruct/mcclean/plsc431/431g.htm
及
http://www.ncbi.nlm.nih.gov/books/bv.fcgi?rid=iga.TOC
http://www.ncbi.nlm.nih.gov/bookshelf/br.fcgi?book=hmg
作業規定:每週指定作業,及平時測驗,累積計分
第1講:課程簡介
遺傳學的主要內涵(The Fields Of Genetics):
傳統或性狀遺傳學(Classical Genetics --based on Mendel's 1st and 2nd laws)、分子遺傳學(Molecular Genetics (based on the Central Dogma of Molecular Genetics))、族群遺傳學(重生物統計)(Evolutionary Genetics --based on the theory of Natural Selection proposed
by Darwin)、遺傳信息(此部份由生物資訊課介紹)
一切生物之結構、行為、繁衍、均受遺傳影響。
吾人生活中接觸的事物,許多均與遺傳有關,如;改良種作物、家畜、疾病(包括癌症)、婚姻生育、老化等。連親子鑒定、犯罪偵測均可用到DNA
fingerprinting 技術。
遺傳的控制,全在於基因。
看動畫:http://life.nctu.edu.tw/~jkhwang/bio-all/MCB/chrom_dna.html
人類有23對46個染色體,其中22對為體染色體,1對為性染色體。
http://www.ncbi.nlm.nih.gov/mapview/map_search.cgi
生物體的生殖,有有性、無性2類。有性生殖可出現基因重新組合,利於演化。
高等生物有性生殖的個體,生物體中有體細胞與生殖細胞2種細胞。體細胞以有絲分裂繁殖,生殖細胞以減數分裂繁殖。
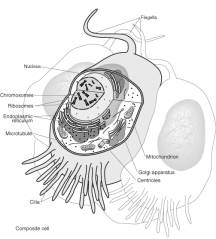
細胞示意圖:
|
|
(http://www.nhgri.nih.gov/DIR/VIP/Glossary/Illustration/cell.html
)
(參考有絲分裂與減數分裂圖;有絲分裂http://esg-www.mit.edu:8001/esgbio/cb/mitosis.html
):
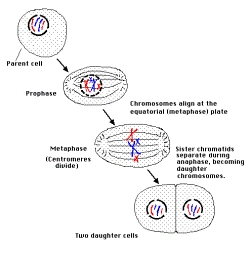
http://esg-www.mit.edu:8001/esgbio/cb/mitosis.html:

在減數分裂時,除聯會交換外,尚有基因排列的分配可造成不同的單套細胞(排列方式有2的n-1次方種,n為染色體的對數)。且聯會交換位置每次不同。故單套配子體種類很多。
看圖說:http://www.csie.ntu.edu.tw/~b4506001/htm/chr-m.htm
(25min)寫心得,並說明有絲、減數分裂之異同、功能比較。
第2講:孟氏遺傳理論
遺傳學之父
Gregor Johann Mendel

孟德爾是農家子弟,出生于現今捷克境內的一個小農莊中 ( Heizendorf, Moravia, Czech Republic),並在當地的學校接受基礎的教育。
西元1843年在他21歲的時候,他加入在Brünn的聖湯馬斯修道院(St. Thomas Monastery)中的奥古斯丁修道會接受神學的訓練,並在1847年的八月被任命為神父。創建於1350年的奥古斯丁修道會與聖湯馬斯修道院在當時曾是一重要的科學研究中心,擁有極多的書籍與植物的實驗栽培園地;尤其是修士們對科學研究的熱中與積極從事科學教育的傳統,啟發了孟德爾對科學與教育的興趣。所以在他成為神父後就開始參與科學的教育,但可惜的是,他並未通過教師的資格考試。為補強他有關自然科學的學院教育與相關的知識,修道院院長同意他到維也納大學就讀。他在維也納大學一共停留了兩年,從1851年到1853年,在期間他修習了有關自然科學與數學等科目,並於1854年返回Bruenn繼續任教。但可惜的是,當他再度參與教師資格考時,他又沒有通過,這次不是因為知識的問題,而是出於健康欠佳。雖然,終其一生他都沒有通過教師的資格考試,但是他在維也納大學所受的教育並沒有白費,尤其是他在數學一科上所受的教育使得他瞭解, 自然界的一切原理原則最簡潔的表明法就數學式的表示法。他也確實將其應用於他的研究中,也因此開創了生物學研究的新視野。
孟德爾自維也納回到Bruenn後就開始在修道院的植物栽培園中進行他著名的碗豆實驗,並首次以數學的方法進行生物學問題的探討。他於1866年將他的成果發表在Bruenn的自然科學史的會刊上,並將他的研究成果寄給相關的科學家。但可惜的是,在他用其他的植物做相同的實驗時,並不能重複的發現他在碗豆雜交實驗中所發現的結果。所以,終其一生,孟德爾所提出的,我們現在稱之為遺傳第一與第二原理的重要發現被埋沒了近三十年,一直到二十世紀初才又被別人再度發現(荷蘭的植物學家 Hugo de Vries , 德國的植物學家 Karl Correns和奧地利的植物學家 E. von Tschermak 分別在他們獨立的植物雜交實驗中發現如同孟德爾碗豆實驗的結果,但不能解釋為什麼,數學的應用找到生物遺傳的基本遺傳法則是孟德爾的最大貢獻)。
在1868年,當孟德爾46歲時,他被選為聖湯馬斯修道院的院長,至此之後,他被繁瑣的修道院事務所佔據。至死,他再也沒有再從事科學方面的相關研究。但終其一生,他念念不忘的就是科學研究,就如同他自己所說的,科學的研究活動帶給他一生最大滿足。
孟德爾死於1884年1月6號。
孟德爾的實驗:
試交(testcross):以1未知基因型的(但通常為顯性)親代,與另1已知為完全隱性的親代雜交,以檢驗未知親代的基因型。
表現型為基因型所控制,同時也受環境的影響。
孟岱爾由1854年起,以碗豆(Pisum
sativum)做實驗,至1865年提出研究成果,卻到死後才為世人所接受。
孟岱爾被稱為遺傳學之父。
他以良好的對照組、詳細的記錄、好運氣的選擇遺傳特性、良好數學基礎、碗豆自花授粉的優點、等而得到成果。
親代(Parental
generation---P)
子代(Filial
generation---F),第1子代---F1,
平滑種皮右鄒種皮2純種雜交,得F1均為平滑,再以F1
x F1 得5474平滑、1850鄒,比率為2.96:1(約3:1)。基於此,他認為,基因均成對出現,在形成配子體時,各自分離(分離律)。且基因之間有顯性與隱性之別。
Results from Mendel's
Experiments
|
Parental
Cross |
F1
Phenotype |
F2
Phenotypic Ratio |
F2
Ratio |
|
Round x
Wrinkled Seed |
Round
|
5474
Round:1850 Wrinkled |
2.96:1
|
|
Yellow x
Green Seeds |
Yellow
|
6022
Yellow:2001 Green |
3.01:1
|
|
Red x
White Flowers |
Red
|
705
Red:224 White |
3.15:1
|
|
Tall x
Dwarf Plants |
Tall
|
l787
Tall:227 Dwarf |
2.84:1
|
Seed Color:
Green and yellow seeds.

Seed Shape:
Wrinkled and Round seeds.

孟岱爾的第2個實驗,就是同時考慮2個不同性狀在雜交後的表現。他以平滑黃種皮SSYY碗豆與鄒皮綠ssyy雜交,得F1均為平黃,而F1
x F1則得9:3:3:1之各種配合比。
|
|
|
Female
Gametes |
|||
|
|
|
GW |
Gw |
gW |
gw |
|
|
GW |
GGWW |
GGWw |
GgWW |
GgWw |
|
Male |
Gw |
GGWw |
GGww |
GgWw |
Ggww |
|
Gametes |
gW |
GgWW |
GgWw |
ggWW |
ggWw |
|
|
gw |
GgWw |
Ggww |
ggWw |
ggww |
由此,他推出獨立分配律:不同性狀的遺傳特性,在形成配子體時各自獨立分配,互不影響。
教活動:試用http://www.geneticswizard.com/f_start_genetics_wizard.asp來算基因機率。(注意:基因要命名)
2個遺傳相關遊戲:http://www.concord.org/biologica/webtest1/web_labs.htm
各組有20min.使用電腦執行上述活動,之後,討論學到什麼?再寫下心得及各設計之優缺點。(25min.)
第3講
Chi Square Test
另外,可以生物統計公式推算:
Chi—square test:
用於檢定2個類別數之關性。
Chi—square
fomular:定義:
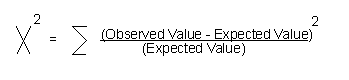
Degrees of freedom (df)
= n-1 where n is the number of classes
Let's test the following data to
determine if it fits a 9:3:3:1 ratio.
|
Observed
Values |
Expected
Values |
|
315
Round, Yellow Seed |
(9/16)(556)
= 312.75 Round, Yellow Seed |
|
108
Round, Green Seed |
(3/16)(556)
= 104.25 Round, Green Seed |
|
101
Wrinkled, Yellow Seed |
(3/16)(556)
= 104.25 Wrinkled, Yellow |
|
32 Wrinkled, Green |
(1/16)(556)
= 34.75 Wrinkled, Green |
|
556
Total Seeds |
556.00 Total Seeds |
![]()
Number of classes
(n) = 4
df
= n-1 =
4-1 = 3
Chi-square value = 0.47
Enter the Chi-Square table at df
= 3 and we see the probability of our chi-square value is greater than 0.90. By
statistical convention, we use the 0.05 probability level as our critical
value. If the calculated chi-square value is less than the 0 .05 value, we
accept the hypothesis. If the value is greater than the value, we reject the
hypothesis. Threrefore, because the calculated chi-square value 0.47 is less than
the 7.82 value, so, we accept the hypothesis that the data fits a 9:3:3:1 ratio.
|
|
Probability |
||||
|
Degrees
of |
0.9 |
0.5 |
0.1 |
0.05 |
0.01 |
|
1 |
0.02 |
0.46 |
2.71 |
3.84 |
6.64 |
|
2 |
0.21 |
1.39 |
4.61 |
5.99 |
9.21 |
|
3 |
0.58 |
2.37 |
6.25 |
7.82 |
11.35 |
|
4 |
1.06 |
3.36 |
7.78 |
9.49 |
13.28 |
|
5 |
1.61 |
4.35 |
9.24 |
11.07 |
15.09 |
Chi-Square
table可參考:http://www.ento.vt.edu/~sharov/PopEcol/tables/chisq.html
或http://www.richland.cc.il.us/james/lecture/m170/tbl-chi.html
或http://www.math.unb.ca/~knight/utility/chitable.html
教學活動:DNA密碼信:http://www.thinkbiotech.com/DNA-o-gram/ (10 min)
1909年,W.L.Johannsen 才以gene來表示孟岱爾的遺傳特性一詞。
特殊基因:
Pleiotropic Effects (多重影的)and
Lethal(致死的) Genes
Epistasis(複基因影響) - the interaction between two or more genes to control a single
phenotype
Modifier genes (修飾基因)- genes that have small quantitative effects on the
level of expression of another gene
第4講
性連遺傳的發現:
1910年Thomas Hunt Morgan 在研究的Drosophila
melanogaster中發現1只公白眼蠅,與一般正常的紅眼果蠅交配,發現白眼為隱性,因為F1全為紅眼,然後他以F1
x F1得F2,中有3470紅眼,782白眼(約18%),不是3:1而是4.44:1。Morgen發現,所有白眼果蠅都是雄性。他因此推論,眼色基因是在X染色體上,雄性為XY,故X上若為白眼,則必為白眼,因Y上無眼色基因。他也發現,此種果蠅生存率較弱。這種與性別相關連的遺傳,為性連遺傳。
就哺乳類和人而言,Y染色體決定男性,無Y就是女性,因為Y染色體上的睪丸決定因數(Testis-Determing
Factor--TDF)促使睪丸之產生,無此則產生卵巢。(在胎兒發育時期中有一些原生細胞等待分化,有TDF則分化為睪丸,無則分化為卵巢)
人類X0
為女性,不能生育、發育不良。
XXY
為男性,不能生育(睪丸發育不完全)。
XXX
為女性,略為低能。
人類對性染色體的多一個少一個有時尚能存活,但體染色體則不能,因為體細胞中染色體凡XX的人均有一個X染色體,雙套中之一(皮膚汗腺的有無為例)緊縮成Barr
Body(Murray Barr發現),故只有一個X染色體發生功能。(在XY的人體細胞中就沒有)。
而這個X是由父方來或母方來,在每個細胞中均不同,自由選擇的。但一但此一細胞選擇其中之一形成Burr
body,則其子細胞均如此不再改。
對果蠅而言,性別決定並非在於Y之有無,而是在於X染色體與體染色體的比率。(autosomes---以A簡示)
果蠅有3對染色體(以A為主),其中一對為性染色體,2對A,因此XY:2A=0.5(雄),
XX:2A=1:1(雌),凡以上此率≧1均為雌性,如果≦0.5為雄性,如果1>此率>0.5
則為兼具二性不完全性性徵且不孕者。
對鳥類、蝴蝶、蛾、及一些魚種,性別的決定與人正好相反,未免與XX系統混淆,而以Z與W方式表示,故ZZ為雄性,而ZW為雌性。
植物中有的雄性花只有花蕊,有的雌性只有雌蕊,有的植物花兼有雄、雌花蕊,為完全花。有些植物有X、Y系統,但多半與果蠅之性別決定方式相同。
對微生物而言,性別並無特別的性染色體,如Saccharomyces
Cerevisiae 只是在一個染色體上有一個基因控制不同的交配型,a或α,稱為MATa或MATα。凡同具MATa的,不相互交配。
也有性別的形成受到環境的影響,如卵孵化的氣溫,或幼體單獨附生則為♀,另一幼體再落在成熟之♀體上則為♂。
將遺傳學原理運用到人的家譜分析,可對結婚生育子女有所諮詢,稱為Genetic
counseling(遺傳諮詢)。可預測子女中發生某一特別遺傳特性的機率。
教活動:找出問題基因。(25min.)http://gslc.genetics.utah.edu/units/disorders/pedigree/mapgene.cfm
|
s+s+ (right) then F1 is right |
ss (left) then F1 is left |
all s+s (right) then all F2 right |
All 3s+_:1 ss then all F3 3 right:1 left |
第11講
量化遺傳
Description
of Quantitative Traits:
These
traits are controlled by multiple genes, each segregating according to Mendel's
laws. These traits can also be affected by the environment to varying degrees.
Here are some images of
quantitative traits in plants:

This image demonstrates the
variation for flower diameter, number of flower parts and the color of the
flower Gaillaridia pilchella. Each trait is controlled by a number of
genes and is a quantitative trait.

The F2
individuals though show a distribution of colors from yellow to reddish orange.
This range o f phenotypes is typcial of quantitative traits.
Statistics
of Quantitative Traits:
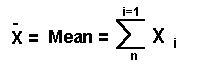
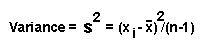
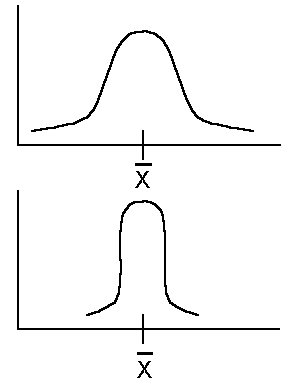

要檢驗1個量化的遺傳值,常用到假設檢定,並通常是為了否定它而設,故稱為Null hypothsis--Ho。若想知道有無不同,則用雙邊檢定,若是比較優劣時,則用單邊檢定。
例:
某校由生中隨機抽取36人進行智力測驗(該測驗之μ平均數=100,σ標準差=16),36之平均為104,請問,在95%顯著水準下,該校可否說生平均智商高於100?
假設
Ho
:μo=100
H1:μo>100
Zo = (104-100)÷(16/√36)=
1.498
而Z0.05 = 1.645,故不可說該校生平均智商高於100
Z Table
可由http://davidmlane.com/hyperstat/z_table.html
網站查。
第12講
演化
Evolutionary
Genetics:
突(mutation)、遷移(migration)、天擇(selection)
教學活動:利用胺基酸比較演化關係(http://www.woodrow.org/teachers/bi/1995/simulation_amino.html
)(30min.)3%
老鼠活動:http://www.explorescience.com/activities/activity_page.cfm?ActivityID=39
(15min)心德2%。
第13講:分子遺傳學
細菌基因的移轉
大腸桿菌(Escherichia
coli)對分子遺傳學的研究十分重要。細菌數量的估計可以玻杆塗平盤法,數菌群來推。
細菌基因的移轉,目前所知有三種方式(本人發現Red
Bridges 尚未能獲公認)
1.Transformation:在細胞外將DNA片段導入細菌中。發生的頻率約在1∕1000,即大約1∕1000的細菌可接受到外來基因。此一部分,將在細胞學、或分子生物學、等課,以實驗方式由同學親自操作,實驗價格昂貴,但技術及經驗很重要,由實驗中可體會出,其中每一個步驟的巧妙與人類智慧的美妙,雖然簡單,但能設想出來是不容易的。
由於基因導入均為片段,相距遠的基因同時導入的可能性就低,(1∕103
×1∕103
=1∕106)大於此數的則表示基因比較接近。
2.Conjugation:不同的細菌間藉由性因數(sex
factor 也稱為fertility factor)所產生的細管,將基因由一方傳至另一方。授予方為F+,接受方為F-。
在E.coli
中,F因數存在於質體中,質體是可以自我繁殖的環形雙股結構的DNA分子,在細菌的染色體之外,此F因數中有若干基因,有些基因導致在細胞表面產生發狀細絲稱為F-pili
或稱為sex-pili ,將F+細胞與F-細胞連接。然後F因數中的一股,由複製原點處裂開,經由細管傳至F-細胞,再在各細胞之單股均複製成完整之F因數。
然後在F因數的細菌中,F因數因聯會而部分插入原細菌之染色體中,此時F因數作用仍在,於是照樣使細菌染色體之一股裂開,並使細菌產生F-pili,而基因因而會傳至接受細菌體內,而這種基因通常都會帶過一些原細菌之基因,而這些基因因有與接受細菌基因相同部分故此較容易交換,故稱為Hfr(high-frequency
recombination)strains。
而Hfr
× F-之後,F-分子並不會成為F+,基因原因為由F+之原點裂開傳送基因,除非全部傳過,否則F+之基因有一部分留在最後,傳不過去,故不能成為F+。
有時F
factor插入細菌的染色體中,又再脫離出來,而脫出來時,帶一些細菌基因出來,成為F',可傳給F-細菌(如其原缺乏此基因時,就會顯出來有了)。
由不同的突變種及不同傳遞時間,可看出基因的距離及先後。
F因數有很多不同的,同一菌中同一種只有一個,不同種的可共存,如插入細菌DNA中,也在不同位置。
3、Transduction:是經由噬菌體將基因由一菌傳送另一菌。此種方式因噬菌體可帶之基因量很少。(通常不到細菌基因之1﹪)
各細菌通常有特別的噬菌體,例如E.coli
有T2、T3、T4、λ等。
T4噬菌體 的繁殖:
1.附著 2.DNA入侵 3. 製造新phages
4. 細菌破裂放出新T4
λ除上述lytic
cycle外,亦有lysogenic pathway,λ插入細菌之染色體固定位置,隨宿主之DNA一同複製,當宿主受UV
light 刺激或營養不足,放射性刺激,…等時,插入之phage
DNA就脫出而開始lytic cycle。噬菌體以何方式將宿主之DNA帶至另一宿主呢?有兩種情形,一為generalized
transduction,這種情形,宿主的任何基因都有可能被帶入另一宿主。另一種方式為specialized
transduction:這種方式為當插入宿主的phage脫出時,帶一點附近的宿主基因走,也留下一點本身的基因在宿主染色體上,因phage插入宿主的位置一定,故所能帶的基因也只限其附近者。
無論上述任一情形,除非兩個基因相當接近,否則不易同時被傳入受體宿主,如發現二個基因markers同時傳入,則此二個基因必十分接近,且頻率愈高者,愈近。
例如A基因與B基因同時傳入有2%的機率,與C基因有50%的機率,而A與C有3%的機率,B與C有0%的機率,則可得推知此三基因排列大致為:
B_____________________A____C_________
基因互補作用(complementation
test):以萬圖說明:
A------------B--------C--------D
A------------B--------C--------D
上2段基因,左邊者,BC失常,右邊CD失常。但所產生的蛋白質可互補。
遺傳學教學活動:耶穌基督的染色體有幾個?(10min)1%
、http://www.bibleplus.org/discoveries/arkintro.htm
第14講
基因的運作
前面一直提及基因,由基因表現出遺傳特性,但到底基因是如何運作的呢?
在1942年由George
Beadle和Edward Tatum做了一個實驗,選出一些變種,在經由各突變種之間與野生突變種的互交,已確定此一種只有一個基因突變。
由此突變,他們提出,一基因一酶的解釋。
但後來發現有的酶由一個以上的基因的產物的形成,故修改為一基因一多月太鏈的說法。
白膚症:Albinism就是由一個染色體上隱性基因突變,使tyrosine不能形成褐色色素melanin,(得此症必為父母雙方所得之基因均此隱性不良基因,因而膚色白,發白,而melanin可吸收紫外線,缺乏,則極為敏感於日照,但是tyrosine形成melanin有兩個步驟,因此,有可能一個人獲得一個不良的基因是在步驟一,另一個在步驟二,則因互補作用,並不會顯出問題。
Tyrosine-->1-->2-->melanin
Sickle-cell
anemia:鐮刀型貧血病:此種症者,在氧氣濃度低時紅血球會形成如鐮刀狀,不如正常紅血球柔韌,造成阻塞微血管,使組織缺氧。這種基因是與正常基因共同顯性的,因此只有一種基因不良者,會有部分正常紅血球,部分不正常,在缺氧情形下,症狀才明顯。此基因是血紅素中的β多月太鍵中一個胺基酸改變,使血紅素的親氧力降低。
A、B、O血型基因,是產生酶,會將不同糖分子加在已有的多糖分子上,這些多糖分子均接在脂類分子上,而形成不同紅血球表面抗元。
教活動---- http://www.biointeractive.org/
2%(15min)
及http://www.woodrow.org/teachers/bi/1995/simulation_amino.html。
(15min)(3%)心得。
DNA的複製
:
DNA複製主要有三程式:
1.DNA
polymerase 在前一個dNTP的3’—OH部分接上新的dNTP(接在5’triphosphate
部分),形成一個新的phosphodiester bond,而必須先有一個Primer﹙一小段的DNA或RNA﹚。
2.DNA模子,使DNA
polymerase可以與一個dNTP來配對。
3.DNA形成方向必定依照5’→3’方向,因DNA
polymerase不會形成3’→5’方向。
3’_________________________________5’
5’-->
E.coli,有三種DNA polymerase,稱為DNA polymeraseI、II及III,I、III均參與DNA之複製,II之功能尚不清楚,I、II及III均有3’→5’endonuclease之功能,可以將在3’的最後一個dNTP消去,這是一種校對的功能,而DNA
polymeraseI也還有5’→3’endonuclease的功能。
DNA的複製均由起始點Origin開始,向前後雙向進行,對DNA量多的生物,Origin有很多對,E.coli只有一對,稱為OriC。
DNA複製的程式:
initiator
Protein附上起始點,隨後DNA helicase附上initiator Protein,使雙股緊黏之DNA打開,隨後DNA
primase,附上形成Primer(這是RNA,所以Primer的基本結構為RNA
polymerase)。
由於DNA只能由5’→3’方向合成,故雙股中的另一股原5’→3’的,就必須等兩股份開後再複製,這種單股DNA片段﹙不可能連續﹚,稱為Okazaki片段。﹙因為Tuneko
Okzaki及其同僚所提出﹚,這些片段,由DNA
polymerase移去Primer,再由DNA ligase接起來。
E.coli至少有下列蛋白質參與DNA複製:
|
Gene |
Function |
|
dnaA,I,P |
Initiation |
|
dnaB,C |
Helicase at oriC |
|
dnaE,N,Q,X,Z |
Subunits of DNA polymerase III |
|
dnaG |
Primase |
|
gyrA,B |
Subunits of gyrase |
|
lig |
Ligase |
|
oriC |
Origin of Replication |
|
polA |
DNA polymerase I |
|
polB |
DNA polymerase II |
|
rep |
Helicase |
|
ssb |
Single-stranded DNA binding
proteins |
就動物細胞而言:
The following table describes
the cell cycle for a hypothetical cell with a 24 hr cycle.
|
Stage
|
Activity
|
Duration
|
|
G1 |
Growth
and increase in cell size |
10 hr |
|
S |
DNA
synthesis |
8 hr |
|
G2 |
Post-DNA
synthesis |
5 hr |
|
M |
Mitosis |
1 hr |
將DNA複製的程式詳細分為幾步驟:
1.雙股螺旋分開(由DNA
helicase為之)。
2.在複製分叉(replication
fork)處之單股DNA由單股附著蛋白質。(Single-strand DNA binding
proteins-->SSB)
3.新的DNA開始合成。
4.合成之DNA延續。
5.滯遲股(lagging
strand)連結。
在複製過程中若有不對的dNTP接上,大多由DNA
polymerase之3’→5’endonuclease功能所除去。
對封閉環狀DNA之複製有二種情形,一為θ型,一為旋轉迴圈型,均可見發生在細菌與濾過性病毒中。
在哺乳類細胞中發現五種DNA
polymerases,稱為α、β、δ、γ及ε。γ只存在粒線體中,其他四種存在核中,α與δ類同DNA
polymeraseⅢ的功能,β與ε用在DNA的修補。而只有DNA
polymerase α有Primase之功能,而粒線體中的DNA
polymeraseγ利用一個DNA
Primase開始複製工作,此DNA Primase是由另一個Primase來合成的,上述五種polymerase均有5’→3’ endonuclease之功能,而只有δε與γ有3’→5’ endonuclease(亦即校正)功能。
我們在很早就提及DNA(染色體)的聯會與複製,到底其詳細過程如何呢?在此,我們略加探討。
由Robin
Holliday所提出的交互聯結模式最為接受:
(http://www.web-books.com/MoBio/Free/Ch8D2.htm
)
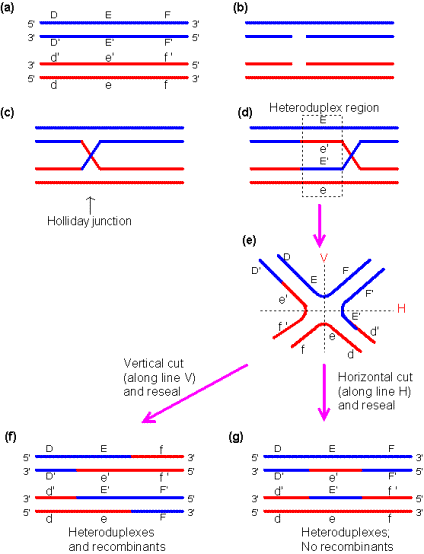
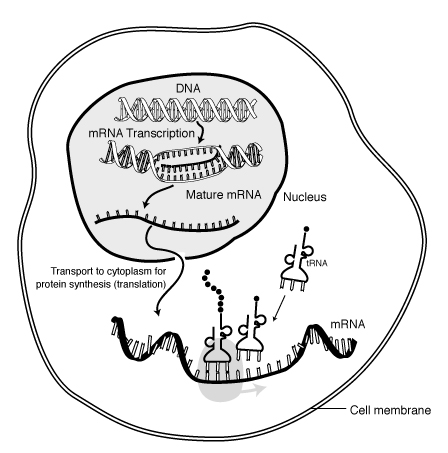
接下來說說基因的運作。基因通常以形成蛋白質來發揮功能,蛋白質又由胺基酸所組成,基因也控制了蛋白質上胺基酸的種類與秩序。DNA首先經由轉錄(Transcription),再經由轉譯(Translation)的程式來形成蛋白質。
轉錄將雙股DNA中一股的資訊改變成單股的RNA的程式。轉譯將傳訊者RNA(mRNA)中的訊息,轉變成製造蛋白質的過程。
在Watson和Crick發表了雙股螺旋型DNA模型理論之後三年(1956),Crick又提出了這種DNA-->RN-->蛋白質的解釋。一個細胞中的DNA並非隨時都在運作,只有部分在產生RNA,這些有在轉錄中的DNA稱為基因的表現。在每個有基因表現的秩序之前,都有一些基因控制碼,作為控制基因表現之用。
教活動:那只牛做的(15min)1%。http://www.flushing.k12.mi.us/fjh/aharvey/mysterybull/
第15講:
遺傳基因宜有三種特性
1、
本身穩定 2、複製正確 3、有變化的可能性(以便演化)。
DNA為遺傳物質的發現:
1928年Frederick
Griffith發現,以平滑菌群(S)(可致病)種之Streptococcus
pneumoniae(以加熱殺死)+粗糙菌群(R)種之S. pneumoniae(活)注入老鼠體內可得貲S種且老鼠會死,因而發現一種可轉殖的遺傳物。
在1944年Oswald
T.avery與Colin M. MacLeod及Maclyn McCarty,他們將S種之菌核酸+蛋白質(核內物)提出,並
1.以蛋白酶處理此提出物,再注入有R種之菌培養液中,可得S活種。
2.以核酸酶RNase處理,再注入有R種之菌培養液中,乃可得S活種。
3.
以核酸酶DNase處理,再注入有R種之菌培養液中,無S活種。
1953年
Alfred D.Hershey及Martha Chase以T2噬菌體分別長於含有P32或S35同位素之培養中,再分別以含P32或S35同位素之噬菌體,長於正常培養液中(含E.coli),待噬菌體附著注入DNA後,以擾動使T2外殼脫去,測得含P32之T2
DNA大量進入E.coli,而外殼部分為S32,說明DNA才是控制遺傳的物質,因為無其他物質進入E.coli促使T2之繁殖也。
目前已知有些濾過性病毒有以RNA為遺傳物質的,例如小兒麻痹病毒polio
viruses。
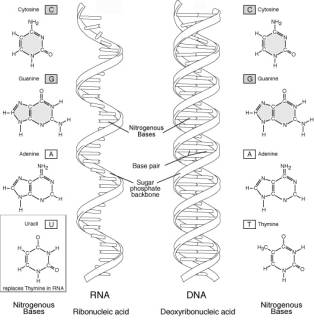
由於它們可被由細胞核中分出且為酸性,故稱為核酸。DNA和RNA均由核甘酸組成,每個核甘酸包含三個部分:
1、五碳糖
2、含氮堿基 3、磷酸根
DNA的五碳糖為去氧核糖,RNA的為核糖
含氮堿基在DNA中有腺嘌呤、鳥糞嘌呤、胸腺嘧啶、胞嘧啶
在RNA中有腺嘌呤、鳥糞嘌呤、尿嘧啶、胞嘧啶
各核甘酸之間由第5碳上的磷酸根與第3碳上的OH部分相結合成鏈。
而DNA是雙股,雙股間由嘌呤與嘧啶之間氫鍵結合而成,由於H鍵形成的方向,故兩股為相反方向,一為5’à3',一為3’à5'。
DNA的結構是由1953James
D. Watson 和Francis H.C. Crick所提出。
他們是依據前人的研究成果整理推測的,在他們之前有Erwin
Chargaff測出DNA中A之數均等於T,G=C。另有X-光繞射的結果,使他們歸納出DAN雙股螺旋形。一般均為右旋(B-form),但也有發現左旋型的Z-form。
故如一股為
5'-TATTCCGA-3'
則另一股為
3'-ATAAGGCT-5'
且他們推出每對堿基間,距離0.34nm(10-9
m),整個360°的一旋為34nm,故一旋中有10對堿基對。
參考:http://www.average.org/~pruss/Nucleosomes/nucDNA.html
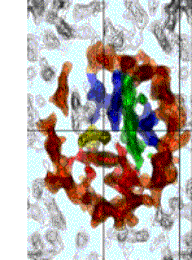
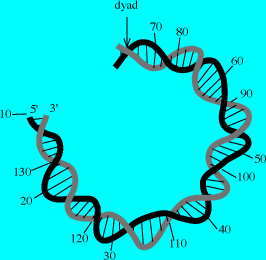
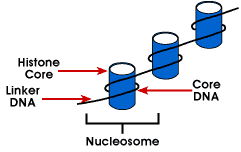
|
|
|
|
(http://www.nhgri.nih.gov/DIR/VIP/Glossary/Illustration/base_pair2.html
)
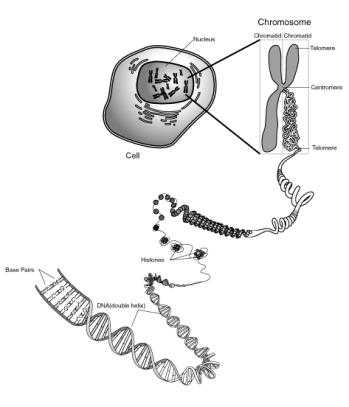
(http://www.nhgri.nih.gov/DIR/VIP/Glossary/Illustration/chromosome.html
)
在瞭解基因如何發生功能前,必須對染色體的結構有所瞭解,細菌的DNA,均為單一的染色體,是高度纏繞方式緊縮。這種方式是由topisomerases所的促成,在E.coli
,topisomeraseII,促成纏緊,而topisomeraseI促成放鬆。
在有核生物的DNA,均與一些變為histones的蛋白質結合,而緊聚在核內。細菌的DNA也與類似的蛋白質結合(在E.coli為HU和H),類似有核生物之DNA與histones之關係。
就人類染色體而言,一般習慣以大小來排列編號,最大的一對編號為1,依次,性染色體則不納入,單獨列X與Y,人體細胞中,含有5.5
×109 base
pairs of DNA(雙套狀態),由於量過大,如無適當結構使之緊縮,必難在核記憶體在,這種緊縮是篇一些蛋白質(histones,H2A,H2B,H3,H4,H1)所促成。形成染色體後,在中心節部分(centromere)為與紡錘絲結合,使細胞分裂時染色體分向兩極的結合外,而染色體兩端有端粒telomere,此存在於每條線狀染色體末端,使染色體不會與別的染色體連在一起,同一種生物的telomeres均有共同的DNA排序。
![]() 人類DNA,依其中排序可分成1.單一,獨有排序 2.中度重複排序
3.高度重複排序。3種。在原核生物,除rRNA基因、及一些tRNAs基因外,均為單一排序。而老鼠的DNA,約70%為單一排序,20%為中度重複排序,10%為高度重複排序。單一或少數重複的基因多為與製造蛋白質有關之基因,中度重複排序(103—104次),之DNA大多無功能,而有功能者多為rRNAs、tRNAs、以及histones基因。(例如人類細胞的160~200重複的rRNA
基因,而且通常這類基因都是一群一群的存在,高度重複排序為重複105à107之多,但多為很短的順序,則未發現有基因存在。
人類DNA,依其中排序可分成1.單一,獨有排序 2.中度重複排序
3.高度重複排序。3種。在原核生物,除rRNA基因、及一些tRNAs基因外,均為單一排序。而老鼠的DNA,約70%為單一排序,20%為中度重複排序,10%為高度重複排序。單一或少數重複的基因多為與製造蛋白質有關之基因,中度重複排序(103—104次),之DNA大多無功能,而有功能者多為rRNAs、tRNAs、以及histones基因。(例如人類細胞的160~200重複的rRNA
基因,而且通常這類基因都是一群一群的存在,高度重複排序為重複105à107之多,但多為很短的順序,則未發現有基因存在。
教學活動:看動畫:http://www.interactive-genetics.ucla.edu/IG_links.htm
寫心得(15min。3%)
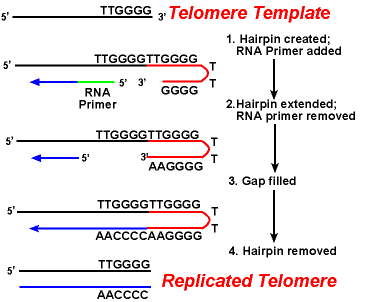
Watson和Crick在提出他們的DNA雙股螺旋模式時,即已指出,如果他們所提的模式是正確的,那DNA的複製就應是雙股份開,每一股作為新DNA的模子,而形成新的兩個雙股DNA,這種模式稱為半保留式。但是這個假設是否成立呢?Mathew
Meselson and Frank Stahl的實驗可說明。
有三種不同模式,他們以E.coli長在15NH4Cl中,得到N
15E.coli,再長於14N液中,如今分散型,則必傘有一代一代的DNA呈現逐步減重情形,而在CsCl離心中呈一條條帶狀。但並未發現此種情形。若為全保留型,則必有各代中均有14N重之帶出現,此情形亦無。只有14N以及介於15N與
14N同之帶出現,故認為是半保留式。
教活動:DNA合成有可能3’à5’方向嗎?若有,對地球上生物有何影?(20min)4%
DNA的合成必須條件:
1、
四種去氧核糖核酸 dATP、 dGTP、dTTP、dCTP、以dNTPs表示之
2、
Mg+2
3、
DNA 模子
4、DNA合成酶(DNA
Polymerase)
DNA複製主要有3程式:
1.DNA
polymerase在前1個dNTP自大3’-OH部份接上新的dNTP
無論原核或真核生物轉錄均由RNA合成酶(RNA
polymerase)的催化。在轉錄的基因之前,雙股螺旋DNA必須先鬆開一部份,以便轉錄,在原核生物,此功能由RNA
polymerase同時擔任,在真核生物則另有一些蛋白質來擔任。
DNA雙股中只有一股轉錄,且轉錄方向必為5’→3’,DNA中3’→5’方向的一股稱為模子股(template
strand)。
RNA
polymerase不要primer,自己可以開始合成RNA,但是沒有校正的功能(Proofreading
abilities),它形成RNA的方式也是以模子上的A配U,G配C的方式。所以,例如:
DNA之模子股若為3’-ATACTGGAC-5’
則RNA為5’-UAUGACCUG-3’
RNA主要有四種:傳信者RNA(messenger
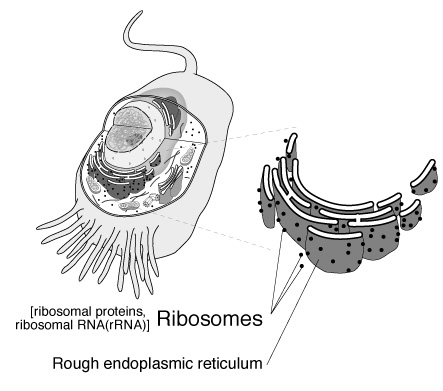
RNA,簡稱mRNA),傳遞者RNA(transfer RNA,簡稱tRNA),核糖體(ribosomal
RNA,簡稱rRNA),以及核內小RNA(small nuclear RNA,簡稱snRNA),因為只有真核生物才有核,故snRNA也只在真核生物才有。
只有mRNA上的訊息會被用來製造蛋白質,製造的場所在核糖體上,核糖體在原核生物是由三個不同之rRNA加上蛋白質所組成,在真核生物則為四個不同的rRNA加上蛋白質所組成。而tRNA則負責將胺基酸帶到核糖體上,配合mRNA之訊息將正確的胺基酸運送過來。SnRNA則與真核細胞mRNA之修飾有關。
RNA剛轉錄成之時,有的並非就是立即有功能的RNA,有的還需要經過化學上的修飾或某些外譯過程才能成為真正有功能的RNA,這點下面還會再詳述。
在細菌(原核生物)只有一種RNA polymerase來轉錄所有的RNA,在真核生物卻有三種不同的RNA
polymerases來轉錄不同的RNA(s),其中RNA polymeaseI只存在於核仁內,負責rRNA之轉錄,包括核糖體中18S,28S,5.8S
RNA(S是Sveberg單位的簡寫,是化合物在等差濃度之蔗糖混合液中離心時沉澱的速率,此值與化合物之大小、形狀都有關係。)RNA
polymerasII只存在於細胞核仁處之核膜外,主要為負責mRNA和一些snRNA之轉錄。RNA
polymeraseIII也只在核仁處之核膜內,負責tRNA、5S rRNA,及一些snRNA之轉錄。
接下來說明原核生物細胞中之轉錄過程:
原核生物的結構性基因(即資訊中是引導製造蛋白質的基因),可分為三類排序,第一類排序是在正式蛋白質暗碼之前,指示基因的起始。第二類排序即正式的蛋白質暗號,即形成mRNA的主體部份。第三類排序為基因結束的指示。第一類排序也稱為起動排列(Promoter),第三排序也稱為終止排序(Terminator)。一般也稱為基因的上游(Upstream),終止排序在基因的下游(Downstream)。
轉錄啟始:
就E.coli而言,在Promoter排序中,有二段DNA排序對轉錄的啟動十分重要,通常它們位在mRNA的第一個排序之前-10與-35附近(-號代表Upstream)。它們通常都有共同類似的排序,在-35附近為5’-TTGAA-3’,在-10附近為TATA為主,故稱為TATA
box(亦有以發現者Pribnow命名為Pribnow box的)。
以E.coli為例:
要轉錄一個完整的RNA
polymerase必須附在Promoter,而且還必須有一個σ因數(Sigma
factor),這σ因數負責辨認Promoter的所在位置,如果沒有σ因數,光只有RNA
polymerase,不能正確的找到起始點,不能有效地開始轉錄。RNA
polymerase與σ因數,首先找到-35的位置附上,在由RNA
polymerase將雙股DNA旋開,至-10的位置,則較緊密的附在整個Promoter位置上,此時RNA
polymerase就開始由+1開始轉錄。由於每個基因的Promoter排序都有所不同,對σ與RNA
polymerase的親和力也就不同,因此每個基因的表現程度也就有所不同。在開始合成mRNA之後,σ因數就由RNA
polymerase脫落。到基因結束時,會有段Terminator排序,及一個蛋白質rho(ρ),凡需要ρ才會終止轉錄的Terminator,稱為ρ-依賴終止排序,有些終止排序為RNA
polymerase本身就可辨識不需ρ之協助,稱為不依賴ρ之終止排序,不依賴ρ之終止排序特性為有回文式的雙向對稱排序,長度約15
~ 20個基對,位置在基因結束前,其後隨之為大約六個AT基對,這種回文式雙向對稱排序可互相構成自我配對,因此在形成轉錄後,此段RNA會形成一個發叉狀配對,在此發叉狀配對之後,接著為一段UUU…造成RNA的轉錄終止並由DNA上脫落,而ρ依賴之終止排序則沒有發狀式及UUU…。在terminator上RNA
polymerase 亦由DNA上脫落。
第16講
就真核生物細胞而言,轉錄製造蛋白質的基因是由RNA
polymeraseII負責,所轉錄而成的是初步mRNA(Pre-mRNA),還要經過修飾與處理才能有功能的mRNA,RNA
polymeraseII也轉錄部分snRNA,至於真核生物的基因控制元則存在于基因的上游與下游,這些控制元(排序)可以與特別的轉錄因數(協助不同的RNA
polymerase來起始)及控制因數(協助活化或抑制基因之轉錄)。依定義,在轉錄起始點之前的控制元(排序)稱為Promoter,真核細胞的Promoter(只對製造蛋白質的基因而言),包括三部分,一為TATA
box,亦稱為Goldberg-Hogness box(依發現者之名命名),一為CAAT排序,一為GC排序,並非所有的真核細胞均有此三者,有的缺其中之一。其中TATA
box的共通排序為5´-TATAAA-3´,此排序大多位在—30左右(-25至-35之間),而CAAT排序之共通排序為5´—GGCCAATCT—3´,此排序位於—80附近,而GC排序則共通排序為5´—GGGCGG—3´,通常在promoter上有多於一個的GC排序區,GC排序的主要功能為協助RNA
polymerase附上接近起始點。另外真核細胞另有促進排序enhancer(s),可促成基因發揮最大的轉錄功能,enhancers通常離開基因起始點有較遠的位置(大於1000基對),而且可以在基因之前或後出現。有些蛋白質負責將此enhancer(s)拉過來並與promoter(s)鄰近以促進基因轉錄。也有抑制基因轉錄的silencer
elements(類似負向enhancers)。
教學活動:做質體DNA:
 \
\
The m-RNA involved in the synthesis of human growth hormone was isolated and found to be:
Study the human DNA and locate the portion that is the gene for growth hormone. Underline or highlight this section.
(http://www.accessexcellence.org/AE/AEPC/WWC/1994/simulation_gene_splicing.html
)
(35min)5%。
RNA
polymeraseII不能自己辨認promoter,需要特別的轉錄因數(Transcription
factors-->TFS)協助,真核細胞的RNA polymerase均有這種需要,故轉錄因數也就命名為TFI-->表示是RNA
polymeraseI的TF,TFII表示是RNA polymeraseII的TF,而每個TFI中又有不只一個,故又加上A、B、C…。
現在來說明一個蛋白質基因的轉錄詳情(為真核細胞,E.coli的前已說明過):
一開始TFIID(亦稱為TATA因數),先附上此基因promoter之TATA排序,同時TFIID也與TIIA結合,又與另一個上游操控(regulatory)因數結合,因此因數又附在enhancer上,將此DNA拉過來,此時RNA
polymeraseII就會因TFIID附在TATA box上,而一起過來附上,此後TFIIB
、TFIIF及TFIIE均過來形成一個轉錄起始複合體,只有其他的轉錄因數附在CAAT及GC
promoter區上。此時RNA polymeraseII就開始轉錄,沿基因前進,而TFIIB和TFIIE就脫離,留下TFIIF,另一個TFIIS在轉錄期間附在RNA
polymeraseII上,而 TFIID、TFIIA及上游操控因數乃留在promoter上,等待另一個RNA
polymeraseII附上以便再開始轉錄工作。
所形成的mRNA,有三部份,在5´端有一段Leader
sequence,在此排序中有一段排序是讓核糖體正確附著以便開始轉錄,此段Leader
sequence本身不被轉譯,Leader sequence之後是真正的coding sequence,就是真正形成蛋白質的部分,之後是trailer
sequence也是不被轉譯的,在原核細胞mRNA轉錄出來即可轉譯,但真核細胞初轉錄出來的RNA還要經過核內的修飾與處理才成為正式mRNA,而且必須送到細胞質內才能轉譯。修飾包括5´及3´兩端及有些introns(無用基因段)之切除與exons(有用基因段)之併合。
在5´部分之修飾是加帽子(cap),稱之為5´
Capping,就是加上一個經由增加甲基的鳥糞嘌呤及2個甲基至RNA的5’兩個基上。這些修飾大約在RNA形成20~30基之時形成,由一個Capping
enzgme來達成。這個5´cap對核糖體附上mRNA極為重要,在3´端則是會被加上50~250個基的腺嘌呤adenine基,因此這個尾巴也被稱為Poly(A)
tail,唯哺乳類的histone mRNAs 沒有Poly(A)
tails。
在mRNA尾端有一對Poly(A)
addition site,在這個site之前約10~30基有一段AAUAAA排序,有一個RNA
endonuclease 會將延續中的mRNA由此site切斷,然後一個Poly(A)
polymerase會將Poly(A) tail加在後面。此Poly(A)
tail可協助mRNA的穩定性。
接下來說明intron(s)之發現,為Philip
Leder等人在研究血紅素基因時,以β-globin
之mRNA與β-globin
之DNA配對,發現有多出之DNA部分,即 intron,introns在形成mRNA時要切除,並將切除之前後mRNA(extons部分)連起來,這種切除,有幾種情形,一種是由snRNAs協助切除,這種切除稱之為mRNA
splicing,而introns通常在5´端均有GU,在3´端均有AG,而snRNA(small
nuclear RNA)及一些蛋白質,構成之snRNP(small nuclear
ribonucleoprotein particles)協助此切除動作,主要有6種snRNAs(U
![]() -U
-U
![]() )及6-10種蛋白質共同為之。
)及6-10種蛋白質共同為之。
以上為蛋白質mRNA的轉錄情形,以下再說明非蛋白質基因的轉錄,包括tRNAs及rRNAs。
rRNA是構成核糖體之RNA,在細菌或真核細胞中,核糖體均分為大小不等之兩部分,每一部份中均有rRNA及蛋白質組合而成,在E.coli中ribosome是30S+50S=>70S,在50S部分有34種不同的蛋白質,一個23S的rRNA,以及一個5S的rRNA,在30S部分,有20種不同的蛋白質,及一個16S的RNA。
在真核細胞的ribosome,是由(以哺乳類為例)40S+60S=>80S,在60S部分有28S,5.8S及5SrRNAs,及約50種蛋白質,在40S部分有18S,rRNA及約35種蛋白質,而在60S中之28S和5.8S
rRNAs是以氫鍵連結的。
第17講:
Central
Dogma of Molecular Genetics:
E.coli的DNA中有7次重複的核糖體RNA,在每一重複中均有16S、23S及5S
rRNAs,介於16S、23S之間及5S之前,亦摻雜有tRNAs基因在內,同時轉錄,再經切開分離。這個核糖體DNA,首先由RNA
polymerase 轉錄為30S Pre-mRNA(P30S),再由RNaseⅢ切為P16S、P23S、及P5S這由特別的酶將這此Pre-mRNA修飾為16S、23S及5S
rRNAs,在rRNAs一形成就立即與核糖體蛋白質相結合而成核糖體。
對真核細胞而言,大多含有大量重複的rRNA(s)基因(18S、5.8S、28S及5S),而18S-5.8S-28S通常排在一起,而且此一組合重複多次(同一片段上重複,再整個基因中亦多次重複此種片斷),而5S之rRNA亦在基因中多次重複。少數生物細胞中的rRNA基因(s)亦含有introns也要經過切除,且切除方式異於前述之mRNA方式。
每個rDNA均由RNA
polymeraseⅠ轉錄成一個大的Pre-rRNA分子(其中含多次重複),將別的ribonucleases來切開此大分子,而5SrRNA是由RNA
polymeraseⅢ轉錄。至於核糖體的蛋白質則是由RNA
polymeraseⅡ轉錄其基因(mRNA)再依轉錄程式形成蛋白質。完整的核糖體是在細胞質內合成。
以下介紹一種introns自我切除的方式,是在一種原生動物Tetrahymena細胞中發現的,它的28S
rRNA含有intron,這種方式不能稱為催化作用,因為它不能還原。(催化劑不能自己產生變化)
5S
rRNA的轉錄起始過程另述。ICR為internal control region(類同Promoter但在基因束之內)。
5S
rRNA通常無intron。
tRNA的轉錄,tRNA基因散佈在基因組(genome)各處,或孕含於tRNA的DNA群(因一個細胞內有至多61種不同的tRNAs),有些真核細胞的tRNA基因含有introns,由特別的endonuclease來切除,再由RNA
ligase將不連之兩段相接。
真核細胞之tRNA(s)基因均由RNA polymeraseⅢ擔任轉錄,真核生物之tRNA基因均有internal control regions為promoter(s),轉錄出的Pre-tRNA均要經過:1.在3´端加上5´-CCA-3´。 2.化學修飾基如pseudouridineΨ、inosine等。此修飾均在核內完成。
(http://www.nhgri.nih.gov/DIR/VIP/Glossary/Illustration/rna.html
)
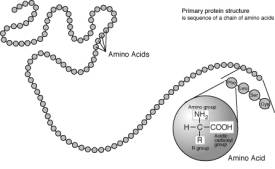
轉錄之後,mRNA的功能在於將基中的密碼轉譯為胺基酸排序。再形成蛋白質。
自然界中生物體大多以20種胺基酸構成蛋白質。胺基酸通常以
R
|
H
![]() N-C-COOH
N-C-COOH
|
H
型式存在,而兩個胺基酸之間以
鍵相連,逐漸形成了多鍵,
R O H
| || |
H
![]() N-C-C-N-C-OOH
N-C-C-N-C-OOH
|
H
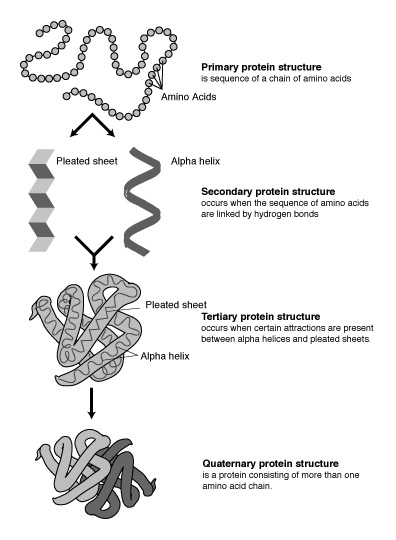
蛋白質有1、2、3、4級結構。
(http://www.nhgri.nih.gov/DIR/VIP/Glossary/Illustration/amino_acid.html
)
(http://www.nhgri.nih.gov/DIR/VIP/Glossary/Illustration/protein.html
在轉譯的時候,mRNA的暗碼是每三個基一組的方式(由A、U、G、C四個基組成的),由於每一個位置均可有四個基可選,故有4×4×4種組合,即64種,但卻只有20個胺基酸,故有些胺基酸可由一個以上的基組標示,何以知道暗碼是三基一組的呢?先是以數量推計,一個基只有4組,不夠20個胺基酸分配,二基一組4×4=16也不只,三基一組有多,但可有彈性,四基一組4×4×4×4=256就太多了,浪費。其他,則為運用致突變藥物促使噬菌體T
4基因突變,由突變之結果比對出暗碼為三位一組。
在得知暗碼為三位一組之後,接下來就是要決定哪個暗碼扣哪個胺基酸,這也是經過各種不同的嘗試與精心設計的實驗而得的,例如,以細胞外蛋白質合成系統(在試管中加入mRNA、tRNA、胺基酸…),將合成的poly(U)
mRNA加入,得全為phenylalaniaes(但poly(G)卻不能成功,因核糖體受poly(G)本身板迭影響不能附著),加入poly(AC)甚至不等量的A與C,使相對比率可得,例如A多於C,結果Asp多於His,則知Asp是兩個A一個C,His是兩個C一個A,或固定的mRNA,如UCUCUCUCUC…,得leu-ser-leu-ser,則可知UCU或CUC暗碼為leu或ser。接著在1964年,由Nirenberg和Philip
Leder發展出來,將三個一組的合成短mRNA放入試管,以20支不同試管加入不同的放射性胺基酸,另19種胺基酸則不做放射性標定,在每一支試管中,加入合成蛋白質所要的核糖體、tRNA(s)及所有64種三基一組的短mRNA,如此一來,在各試管中的核糖體、短mRNA,和與帶有正確胺基酸的tRNA結合,將各試管分別倒至不同之濾紙上,凡未能與核糖體、tRNA、mRNA三者結合者,均會流走,只剩三者結合者,如此就可分析出哪一個三基因組是哪一個胺基酸暗碼了。暗碼均以5´→3´方式寫。
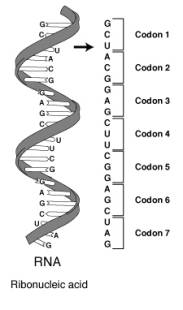
歸納之,mRNA暗碼的特性:
1. 三基一組。
2. 連續無間隔。
3. 不重迭。
4. 幾乎所有細胞均同。
5.
暗碼的第三碼有彈性(每個胺基酸有時有不只一個暗碼),尤其是當前兩碼相同,而第三碼是U或C,幾乎都是扣著同一個胺基酸,或前兩碼相同,而第三碼為G或C,也都幾乎是扣同一個胺基酸。
6. 暗碼有起始碼與終止碼(AUG通常是起始碼,GUG也有但很少用),(且原核、真核細胞均以AUG起始)(起始碼本身也扣胺基酸,但終止碼則不扣UGA、UAG、UAA三碼為終止碼)。
7. tRNA上的反扣碼(anticodon)也有彈性,即原來必須61個tRNA(s)來辨認61個暗碼,卻因tRNA之反扣碼上的基因可以與非固定配對的基配對,而產生彈性。(如講義之表,稱Wobble)
在轉譯時,mRNA是依5´→3´方向轉譯的,而蛋白質由N端向C端延長。正確的安基酸排列有賴於暗碼(mRNA)與反扣碼(tRNA)的配對及胺基酸附在正確的tRNA上,轉譯的三步驟在原核與真核細胞均大致相同。
教學活動:家譜研究題:http://www.sc2000.net/%7Eczaremba/labs/pedigree.html
(15min)2%
第18講
正確的胺基酸(a.a)先由一種Aminoaiyl-tRNA
synthetase促成附在tRNA上,因有20種不同的a.a(s),故也有20種不同的之Aminoaiyl-tRNA
synthetases。同時由於暗碼的彈性,故所有可帶同一種a.a的tRNA(s)均由同一種Aminoaiyl-tRNA
synthetase來執行。其過程如講義說明。
在原核與真核細胞,轉譯均由AUG起始,AUG也扣methionine,故所有的蛋白質均以met起始,有些後來被切除。在原核生物,起始點的met被修飾由formylmethionine(簡寫fMet)表成為起始胺基酸,並由一個特別的tRNA,成為tRNA.fMet來帶,這個tRNA的反扣碼為
5´-CAU-3´與5´-AUG-3´相配。帶有fMet之這個tRNA,可寫成fMet-tRNA.fMet,此tRNA只用在起始點,在起始點外的AUG均由其他的tRNA來帶fMet過來。稱為tRNA.fMet,但卻均由同一個aminoaiyl-tRNA
synthetase來促成胺基酸的附上。tRNA.Met與tRNA.fMet分子由不同的基因所暗扣並有不同的排序。
在真核生物AUG也是起始碼,但N端的methionine並無
group,但起始的tRNA.Met與非起始的tRNA不同。
無論原核或真核細胞均是核糖體的小半部分先附上mRNA,加上其他的蛋白質協助,大半部分再附上,形成一個起始複合體。再開始蛋白質的合成。
在Prokaryotes除AUG碼外,在mRNA的5´端,在AUG碼之前還要有一段排序,稱為Shine-Dalgarno排序,使核糖體能附上,此段排序為5´-UAAGGAGG-3´,這與核糖體上小部份之16SrRNA的一段5´-CCUCC-3´正可配對。
(http://www.nhgri.nih.gov/DIR/VIP/Glossary/Illustration/codon.html
)
(http://www.nhgri.nih.gov/DIR/VIP/Glossary/Illustration/mrna.html
)
(http://www.nhgri.nih.gov/DIR/VIP/Glossary/Illustration/ribosomes.html
)
在E.coli
三個initiation factors,IF1、IF2及IF3,附上30S
rifosomal sufunit,及一個分子的GTP(guanosine tiphosphate),然後fMet-tRNA.fMet及mRNA就附在
30S-IF-GTP的複合體上,之後IF3脫落,50S附上,GTP
水解而IF1及 IF2脫落,這時70S核糖體已形成,並以附上mRNA的正確位置,70S
rifosome有兩處可供aminoaryl-tRNAs填入之處,一為peptidyl(p)一為aminoaryl(A)處,而
fMet-tRNA.fMet是進入P處,並扣上mRNA。
真核細胞則以真核細胞的initiator
fartor eIF4A,就認出mRNA的5´端Cap附上,隨mRNA移動來找尋AUG暗碼,一但找到40S就附上,60S也就附上形成80S
,其中Met-tRNA.Met就落入P處。
接下來是轉譯的持續,包括三個步驟,1為Amionaryl-tRNA附進核糖體,2為肽鍵之形成,3為核糖體延mRNA一次一個暗碼的移動。
轉譯初起時,以E.coli為例,fMet-tRNA.fMet以氫鍵與AUG碼結合於P處,以後另一個Amionaryl-tRNA由蛋白質延長因數(elongation
factor)EF-TU-EF-TS複合體,再分開為EF-TU-GTP與EF-TS,另一個Amionaryl-tRNA再與EF-TU-GTP結合再開始迴圈工作。
肽鍵的形成是由peptidyl
transferase 促成,而此功能是由23SrRNA所擔任,一但肽鍵形成,一個無胺基酸的tRNA就由P處脫出。此時另一個elongation
factor EF-G 形成EF-G-GTP複合體並附上核糖體,使核糖體向前移一暗碼,而失去a.a的tRNA亦由P處脫出。其後EF-G也在消耗一個GTP的情況下釋出,在核糖體移位之時,此peptidyl-tRNA
自然成為在P位置。此時A位已空出,另一個Amionaryl-tRNA具有正確的反扣碼者即可再進入。此種程式重複直至停止碼為止。在真核細胞其程式大致相同,只是elongation
factor數量與性質不同。
無論原核或真核細胞,在一個ribosome離開起始碼後,另一個ribosome就可進入,形成polyribosome(或
polysome)現象。
當遇到UAG、UAA或UGA(無論原核或真核細胞均同)碼時,在termination
factors或release factors的協助下,核糖體辨識到終止碼,而後轉譯終止。
E.coli
有3個RFs→RF1、RF2﹠RF3
RF1可辨識UAA及UAG
RF2可辨識UAA及UGA
RF3不辨識任何終止碼,只促成終止行動
在真核細胞卻只有一個eRF,可辨識三個終止碼
由RF(s)所促成的終止行動包括:
1. 使多肽鍵在Psite的tRNA上脫去。
2. 使tRNA由核糖體上脫去,並促使核糖體的二半部份分離,離開mRNA。
蛋白質製造出來,要送到需要之處,這個運送也受基因控制,
通常要分別運送的蛋白質,是前端N-端,都會多一段訊號排序(signal
sequence),此段與signal recognition partide(SRP)結合,在附至目標區上的docking
protein,使此蛋白質送入,(見講義)。之後此signal sequence不用時會被酶切去,在真核細胞還在高氏體可協助運送蛋白質並修飾之。
教學活動:線上DNA
fingerpring Lab :
http://www.pbs.org/wgbh/nova/sheppard/labwave.html
(15min)心得3%
第19講
接下來說明遺傳工程技術:
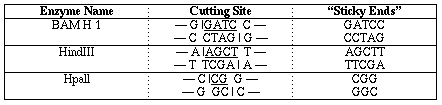
這個技術是由限制酶的使用所發展出來。(Restriction
Enzymes或稱Restriction endomuleases),這種酶通常可在某一特別DNA排序中將雙股DNA切斷,這種酶原均由細菌中分離出來,它們原為保護細菌免受來病毒之侵害,凡細菌之DNA中有相同排序處均自我修飾過。
限制酶依它們分離出的菌種命名,頭三個字以斜體字或加底線方式,加上一個羅馬數字,有時再附加字母以特別表明分離出它的菌種系(Strain),例如:EcoR1是由E.coli
Strain R13來的,HindⅢ是由Haeonophilus
inqluenzae Strain Rd來的,它切DAN大致均由3´-OH
與PO相接處,故留下的均為5´-PO及3´-OH部份,例如EcoR1,切
↓
5´-GAATTC-3´
3´-CTTAAG-5´ (回文結構)
|
切斷處
也有如HaeIII的會切成平頭DNA雙股而非錯開式。
DNA ligase則可將錯開式的各股又連接起來。(平頭式亦可但較不易成功)
在遺傳工程技術中最常用的基因導體(Cloning
vectors),首推Plasmids,小的雙股環型DNA,存於細菌中,一個Plasmid基因導體,有下列條件:
1. 必須有Ori排序以便複製
2.
有可篩選的特性(Selectable
/marker),如Ampr,以便始有此Plasmid的細菌與沒有的,可以區別
3. 單一的限制酶切斷點
也有人設計出一個PUC19(PUCK-19),其中有多重插入點,如講義說明:當有基因插入插入點後,以DNA
ligase連接,再以transformation法送入細菌,在測出可否測出白色菌群(Blue表示乃有Lac
Z),長在含Amp之平板上,則可選出所要的細菌。
目前已有一些基因導體可適合多種用途(既可用於細菌,亦可用於動、植物細胞)
也有用噬菌體為導體的,如λ將lysogeny部份去掉,在λ之兩端有重要基因(切除部份不必要基因,不重要基因可取代,並有單一的限制酶切點),以插入基因,這樣的新結構因兩端基因量占去位置,最多可插入15kb的外來基因。
基因圖書館(資料庫genomic
library)之建立:
將生物基因切成一段段,放入導體中,以便未來運用,如將人的基因依不同染色體,可將分裂中的細胞以一種螢光染色劑染色,再將細胞漲裂,以雷射光分辨不同染色體,並可自動分離之,再將每個染色體分切,裝入導體,以後要用時再以探針去找。(如講義說明)
Complementary
DNA(cDNA)之製作:
由mRNA用reverse
transcriptase來做,如講義說明
而細胞中的mRNA如何取得呢?以細胞中的RNA(s)通過oligo(dT)column,再加熱column(以熱buffer通過)使脫下取得之。
對Blunt
ends 的DNA也可做遺傳工程:
DS
NDA如:
T4
ligase + 5´-GAATTC-3´
3´-CTTAAG-5´
↓
↓
![]() 5’ GGATCC --------------GGATCC
3’
5’ GGATCC --------------GGATCC
3’
![]() 3’
CCTAGG
--------------CCTAGG 5’
3’
CCTAGG
--------------CCTAGG 5’
↑
↓
↑
以Bμm HI切之
形成
![]() 5’
GATCC
G 3’
5’
GATCC
G 3’
![]() 3’G
CCTAG 5’
3’G
CCTAG 5’
則成為sticky ends 了!
對cDNA,製成後必須使其式為expressing
vector,即前有promoter,後有terminator蛋白質製造出之後,將細胞裂開於nitroccellulose上,加上獨對此蛋白質之antibody,而此antibody上則加上ridio
label或螢光染色。
Restrictroin
mapping:以不同限制鋂切DNA,再電泳以DNA size標準算出各段長度,再以智力拼圖為之,如講義之例說明之。
在限制切過的DNA中找出有無所要的或類似的已知基因,可用southern
Bloting方式測之。(如另說明)
*m5' - G A A T T C - 3' (*Remember the enzyme will not 3' - C T T A A G - 5' cut if the 3' A is methylated.) m*
DNA排序分析:(DNA
sequence analysis)
目前有兩種方法,一種Maxam-Gilbert
sequencing,1另1為Dideoxy sequencing,目前後者較常用。
其次介紹聚合鋂連鎖反應(Polymese
Chain Reactior PCR):以電影說明如下
教活動:http://www.dnalc.org/resources/BiologyAnimationLibrary.htm
(15min)心得3%。
DNA
fingerprinting(另依本人論文講解)。聽後心得5%。
運用遺傳工程技術可協助早期偵測胎兒之染色體正常有無特殊遺傳疾病。
另有DNA分析的(RFLPs)運用很廣,由犯罪學、親人找尋、分類學…..均可用到。
基因治療法:將有缺乏或不對之基因的細胞注入好基因,使其產生好蛋白。
第19講
人類基因組(另依本人論文講解)。聽後心得5%。
人類基因計算:
人有3.2
×109bp的基因,約3—14萬個基因,希望在今年底前將人之DNA全數排序出來。
至於用遺傳工程技術於生產商業用途的蛋白質,農業改良更是太多。以Tomatoes易爛為例,已有人將此易熟基因(PG)倒裝
將CG….GA倒裝回去,產生的mRNA正好是正常,PG的反mRNA(anti
sense mRNA)就會中和掉與之成雙股而不生作用,使Tomatoes不易早熟,不易爛。
第20講:
以下說說細菌與噬菌體的基因控制:
細胞中有些基因受環境因素影響,有些不受而經常運作者稱為(constitutie
genes)
有些基因是可引導的(inducible),此導引物稱為導引子(induer)轉錄這些可導引基因的控制在於此DNA前會有一段contolling
site。
下麵以E.coli的Lactose
operon說明:
Lactose是由glucose+galactose合成,E.coli會先用glucose,無時才用Lactose。
參考:
LacI
protein: http://www.rcsb.org/pdb/cgi/explore.cgi?pid=27191959383417&page=0&pdbId=1LBG
看電影:http://www.dartmouth.edu/~cbbc/courses/movies/LacOperon.html
參考:http://www.acsu.buffalo.edu/~jbarnard/LacOperonII.html
及http://140.116.60.1/mdlai/Handout/transcription-regulation-med-99/sld001.htm
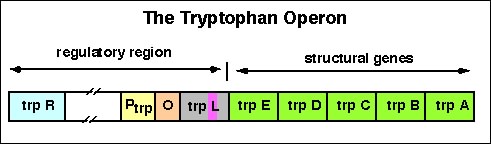
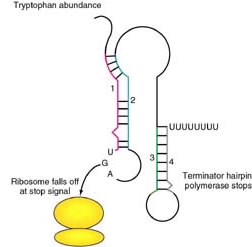
Tryptophan
operon: http://www.blc.arizona.edu/marty/411/Modules/trpop.html
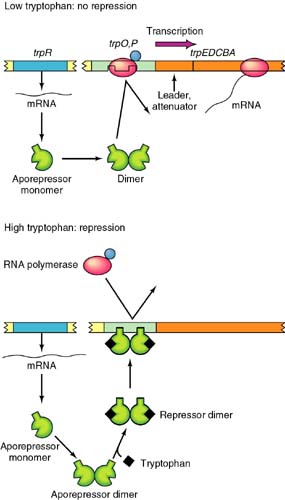
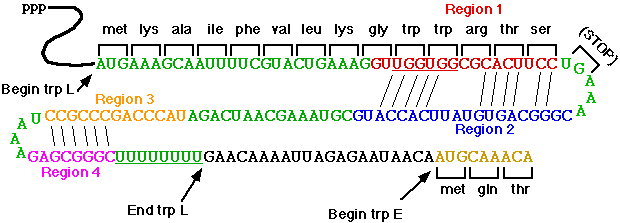
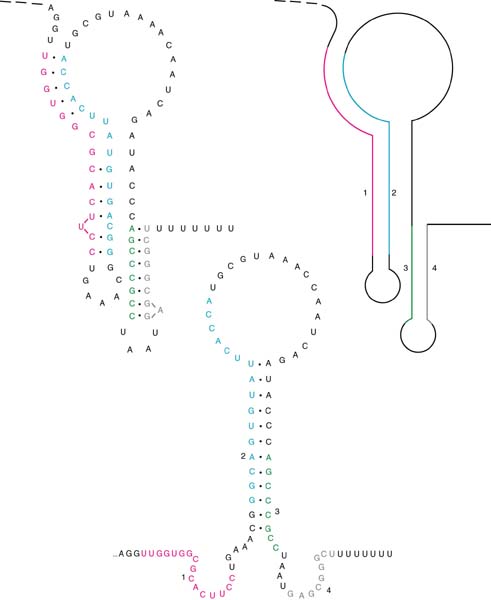
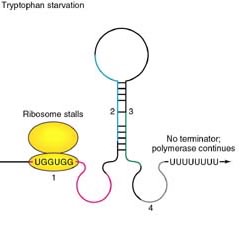
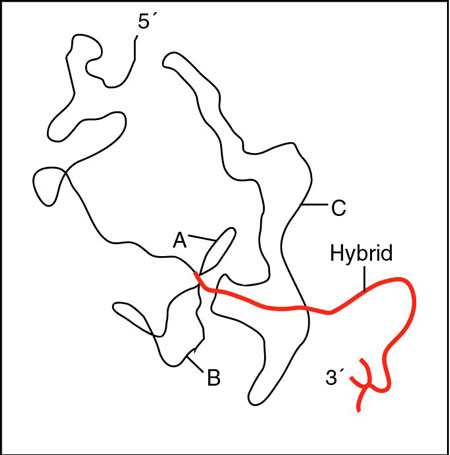
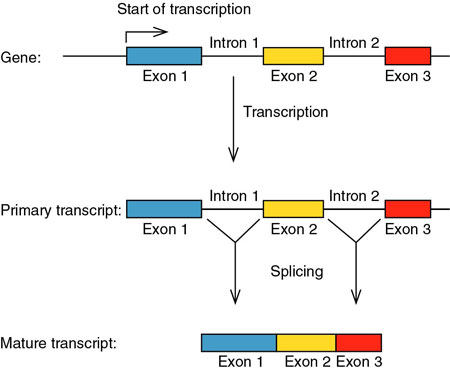
RNA
splicing: http://www.blc.arizona.edu/marty/411/Modules/mod15.html

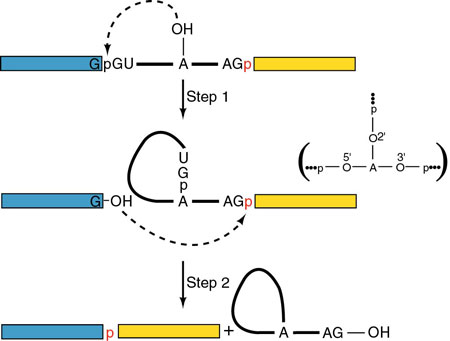
Splicing signals have been identified.
The signal identified for higher eukaryotes is:
5' - AG|GUAAGU – intron – YNCURAC
– YnNAG|G - 3'
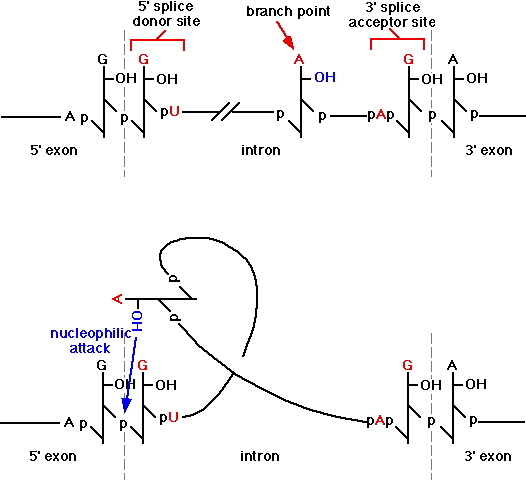
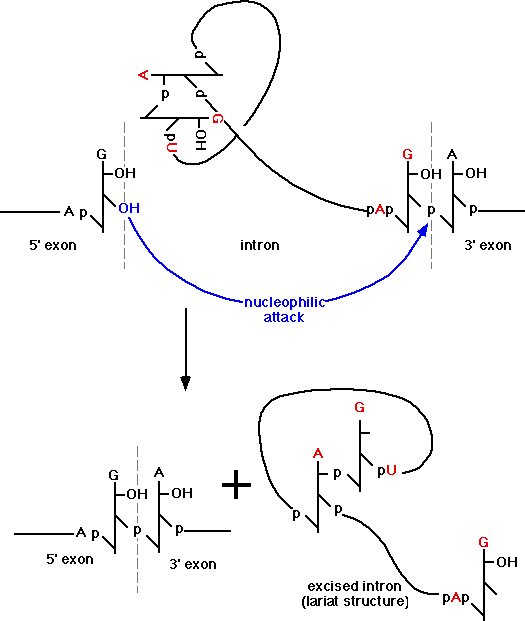
心得5%
第21講:抗體基因
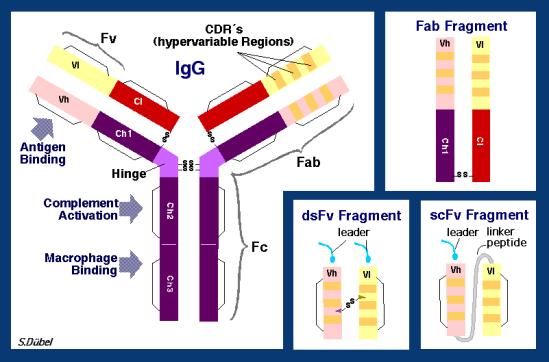
(http://www.mgen.uni-heidelberg.de/SD/IgG_Fv.GIF )
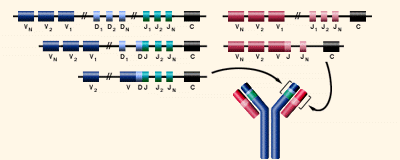
(http://www.biology.arizona.edu/immunology/tutorials/immunology/05t.html )
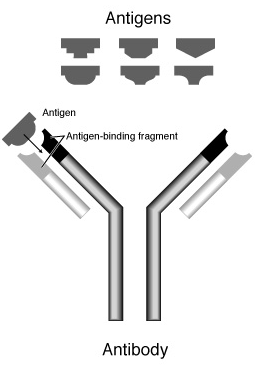
(http://www.nhgri.nih.gov/DIR/VIP/Glossary/Illustration/antibody.html )
第22講:胚胎發生
參考:http://users.rcn.com/jkimball.ma.ultranet/BiologyPages/H/HomeoboxGenes.html
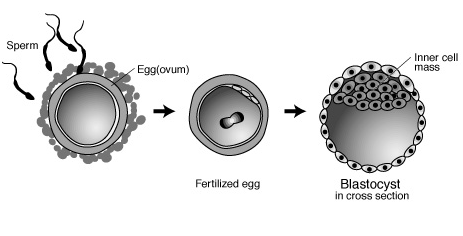
(http://www.nhgri.nih.gov/DIR/VIP/Glossary/Illustration/blastocyst.html
)
心得4%。
第23講:突變
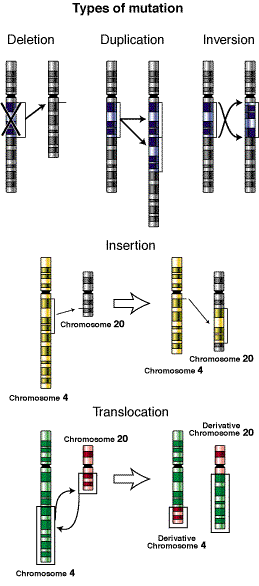
(http://www.nhgri.nih.gov/DIR/VIP/Glossary/Illustration/mutation.html
)
第24講
質體載體(另依本人論文講解),聽後心得5%。
第25講
斡細胞療法(另依本人論文講解)。心得5%。
第26講
生物技術與人類未來(另依本人論文講解)。心得5%。

{kind=link}